
搜索标签
标题关键词基因工程 ; 细胞工程 ; 酶工程
技术功效大幅提高了人溶菌酶基因mRNA稳定性; 密码子优化大幅提高了重组人溶菌酶表达量; 通过检测荧光素酶可以实现快速、准确地评定密码子优化效果; 通过检测荧光素酶可以实现快速、准确地评定密码子优化效果; 显著提高了人溶菌酶在牛乳腺上皮细胞及成纤维细胞中表达; 提高人溶菌酶编码基因中对应氨基酸牛优势乳蛋白高频密码子; 提高人溶菌酶编码基因中对应氨基酸牛优势乳蛋白高频密码子; 有利于后期通过工业化生产重组人溶菌酶; 可以快速、准确的对密码子优化效果进行评价; 显著提高了人溶菌酶在牛乳腺上皮细胞及成纤维细胞中表达; 通过检测荧光素酶可以实现快速、准确地评定密码子优化效果; 减少低频密码子
应用场景无
技术问题无
IPCC12N15/56
技术参数无
著录项
申请号CN201810257256.9
公开号CN108624606A
申请(专利权)人西北农林科技大学
主分类号C12N15/56
地址712100 陕西省西安市杨凌示范区邰城路3号
代理机构西安恒泰知识产权代理事务所
申请日20180327
公开日20181009
发明人成功;昝林森;王力
分类号C12N15/56;C12N15/62;C12N15/85
国省代码CN61
代理人李郑建
摘要
本发明涉及一种密码子优化的重组人溶菌酶基因及其应用,该密码子优化的重组人溶菌酶基因是通过比较牛乳腺优势乳蛋白高频密码子和低频密码子对人溶菌酶密码子进行全局优化(Lyzop)和翻译起始后23位密码子进行优化(Lyzop23)。优化后的密码子显著提高了人溶菌酶mRNA稳定性及在牛乳腺细胞中mRNA表达量。通过蛋白免疫印迹实验证明,密码子优化的重组人溶菌酶基因在牛乳腺细胞中表达量是野生型的2.18倍。为今后细胞生物反应器以及牛乳腺生物反应器高效生产重组人溶菌酶提供了工具基因。同时,将密码子优化的重组人溶菌酶基因与荧光素酶融合表达,通过分析荧光素酶表达量,实现密码子优化效果的快速、准确评价,为今后开展基因密码子优化提供了新的可靠评价方法。
权利要求书
1.一种密码子优化的重组人溶菌酶基因,其特征在于,该密码子优化的重组人溶菌酶基因是Lyzop和Lyzop23,它们的核苷酸序列为:其中,斜体加粗部分表示重组人溶菌酶基因按照牛乳蛋白偏好密码子进行设计优化后的密码子。
2.权利要求1所述的密码子优化的重组人溶菌酶基因在乳腺生物反应器的应用。
3.如权利要求2所述的应用,其特征在于,所述的密码子优化的重组人溶菌酶基因与PGL3-control载体经NcoI实现与荧光素酶融合的表达载体。
4.如权利要求2所述的应用,其特征在于,所述的密码子优化的重组人溶菌酶基因与荧光素酶融合评估密码子优化效果。
[收起]
说明书
技术领域
本发明属于分子生物学、细胞工程技术领域,具体地涉及一种密码子优化的重组人溶菌酶基因及其应用。
背景技术
溶菌酶又称胞壁质酶或N-乙酰胞壁质聚糖水解酶,是一种能水解致病菌中粘多糖的碱性酶,具有广谱抗菌、消炎、抗病毒等多种功效。人溶菌酶作为天然蛋白,在人体内体液、细胞及组织器官中广泛分布,发挥重要抗菌、抗病毒、调节机体免疫等作用。目前,人溶菌酶在临床疾病治疗、乳制品添加剂、食品防腐等方面具有广泛应用。然而,传统方法从母乳、胎盘或尿液分离提取很难达到工业化生产要求。利用细胞生物反应器或乳腺生物反应器相比细菌、酵母等生物反应器具有稳定性好,重组蛋白活性高等优点,但普遍存在着重组人溶菌酶表达量低的问题,不利于下一步工业化生产。
组成蛋白质的氨基酸有20种,而编码氨基酸的密码子有64种,每个氨基酸至少对应1种密码子,最多可以有一种氨基酸对应6种密码子(简并密码子)。不同生物,甚至同种生物不同的蛋白质编码基因,对简并密码子使用频率并不相同,具有一定的偏爱性,其中使用频率高密码子称为最佳密码子(optimal codons),而那些很少被利用的密码子称为稀有或低频密码子。在生物反应器研究过程中,外源基因含有宿主细胞基因使用的低频密码子或者偏好密码子不一致情况下,往往会影响外源基因的mRNA的稳定性和翻译效率,造成重组蛋白表达量低。前期研究结果发现,重组人溶菌酶在转基因羊、牛乳腺生物反应器中表达量相比其他重组乳蛋白表达均不理想,造成后续提纯生产成本高。
发明内容
本发明的目的在于,提供一种密码子优化的重组人溶菌酶基因,以及利用该密码子优化的重组人溶菌酶基因在乳腺生物反应器的应用。
为了实现上述任务,本发明采取如下的技术解决方案:
一种密码子优化的重组人溶菌酶基因,其特征在于,该密码子优化的重组人溶菌酶基因是Lyzop和Lyzop23,它们的核苷酸序列为:
GCCTAGCAAACTGGATGTGTTTGGCCAAATGGGAGAGTGGTTACAACACACGAGCTACAAACTACAATGCTGGAGACAGAAGCACTGATTATGGGATATTTCAGATCAATAGCCGCTACTGGTGTAATGATGGCAAAACCCCAGGAGCAGTTAATGCCTGTCATTTATCCTGCAGTGCTTTGCTGCAAGATAACATCGCTGATGCTGTAGCTTGTGCAAAGAGGGTTGTCCGTGATCCACAAGGCATTAGAGCATGGGTGGCATGGAGAAATCGTTGTCAAAACAGAGATGTCCGTCAGTATGTTCAAGGTTGTGGAGTGTAA;
其中,斜体加粗部分表示重组人溶菌酶基因按照牛乳蛋白偏好密码子进行设计优化后的密码子。
根据申请人的实验表明,上述密码子优化的重组人溶菌酶基因能够在乳腺生物反应器的应用。
进一步地,所述的密码子优化的重组人溶菌酶基因与PGL3-control载体经 NcoI酶切位点连接实现与荧光素酶融合的表达载体。
所述的密码子优化的重组人溶菌酶基因与荧光素酶融合评估密码子优化效果。
本发明的密码子优化的重组人溶菌酶基因,与现有技术相比,具有以下优点:
(1)选取了牛乳中7个高丰度表达的乳蛋白基因进行密码子偏好性分析,牛乳蛋白基因对CTG(2.71)、AGG(2.32)、GCC(1.73)、GTG(1.68)、ATC (1.64)等5个密码子具有明显的偏好性(RSCU>1.5),为高频密码子。而TTA (0.12)、ATA(0.18)、TCG(0.19)、CTA(0.25)、CCG(0.26)、GCG(0.45)、 ACG(0.48)、TAG(0.43)等8个密码子在牛乳蛋白基因中使用频率较低(RSCU<0.5),为低频密码子。为开展牛乳腺生物反应器密码子优化提供理论和技术依据。
(2)牛乳蛋白密码子第三位碱基偏好以GC结尾,人溶菌酶偏好以AT结尾。依据计算、分析获得的牛乳腺中乳蛋白高频密码子(使用频率较高)及低频密码子(稀有密码子)对人溶菌酶密码子进行优化,提高人溶菌酶编码基因中对应氨基酸牛优势乳蛋白高频密码子,减少低频密码子。比较发现,在不改变人溶菌酶蛋白质序列的基础上,大幅提高了人溶菌酶基因mRNA稳定性。同时,通过qPCR在牛乳腺上皮细胞、成纤维细胞、小鼠C127细胞及Western-blot 在BMEC细胞中分析证明,密码子优化大幅提高了重组人溶菌酶表达量,有利于后期通过工业化生产重组人溶菌酶。
(3)人溶菌酶基因编码片段与PGL3-control载体通过NcoI酶切位点连接,实现人溶菌酶基因与荧光素酶基因融合表达。细胞转染后24小时即可通过检测细胞中萤火虫荧光素酶(Luc)表达量来评判目的基因密码子优化效果。同时共转染Prl-tk,载体中海肾荧光素酶(Rluc)可作为内参校准转染效率、细胞数量之间差异,可以快速、准确的对密码子优化效果进行评价。
通过密码子优化,显著提高了人溶菌酶在牛乳腺上皮细胞及成纤维细胞中表达,为今后利用细胞生物反应器及牛乳腺生物反应器生产人溶菌酶提供了基因载体。同时,利用密码子优化的重组人溶菌酶基因与荧光素酶基因融合,通过检测荧光素酶可以实现快速、准确地评定密码子优化效果,为密码子优化效果评价提供了一种新的方法。
附图说明
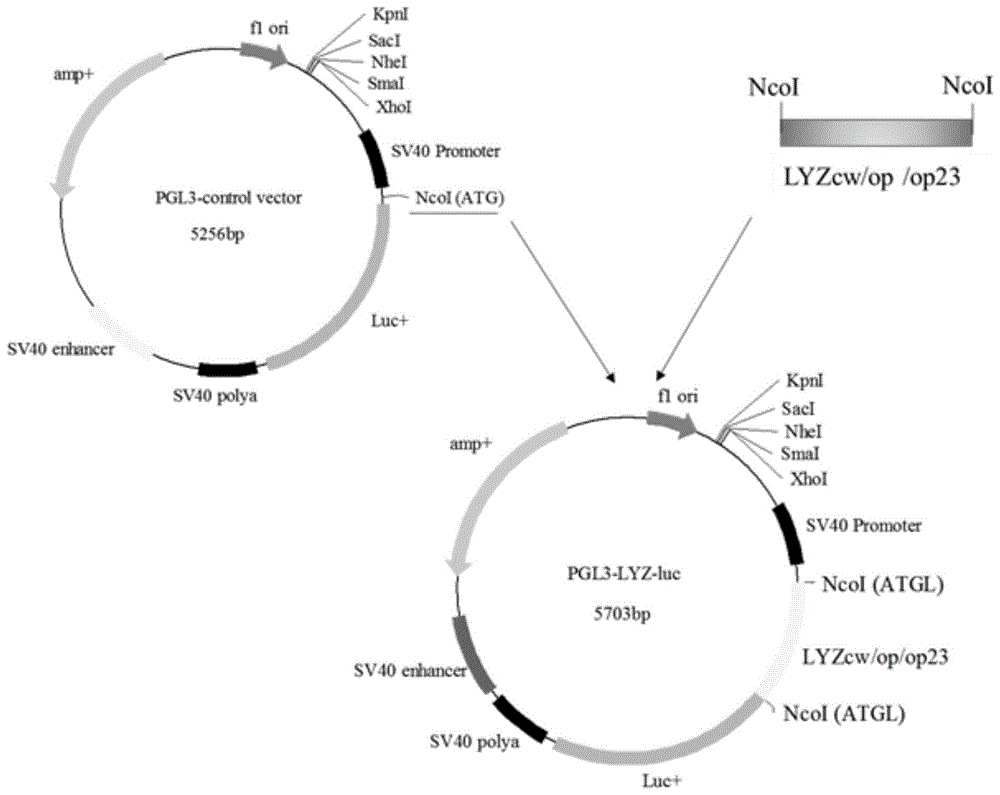
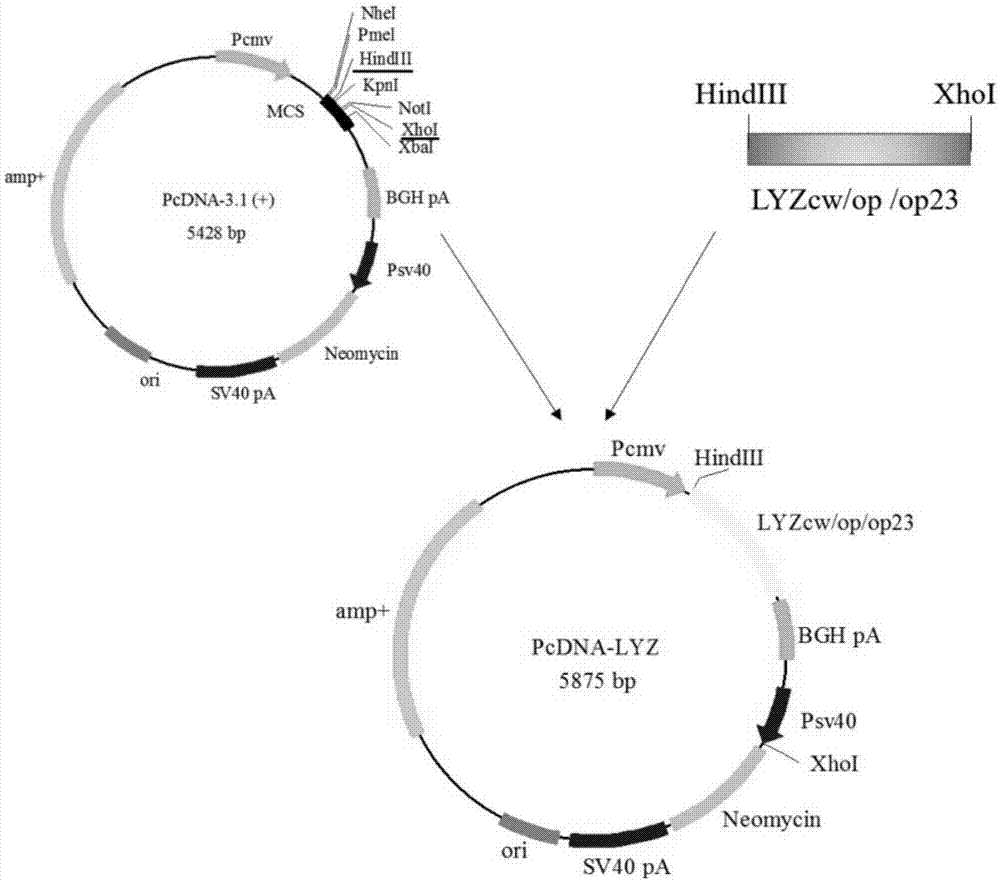
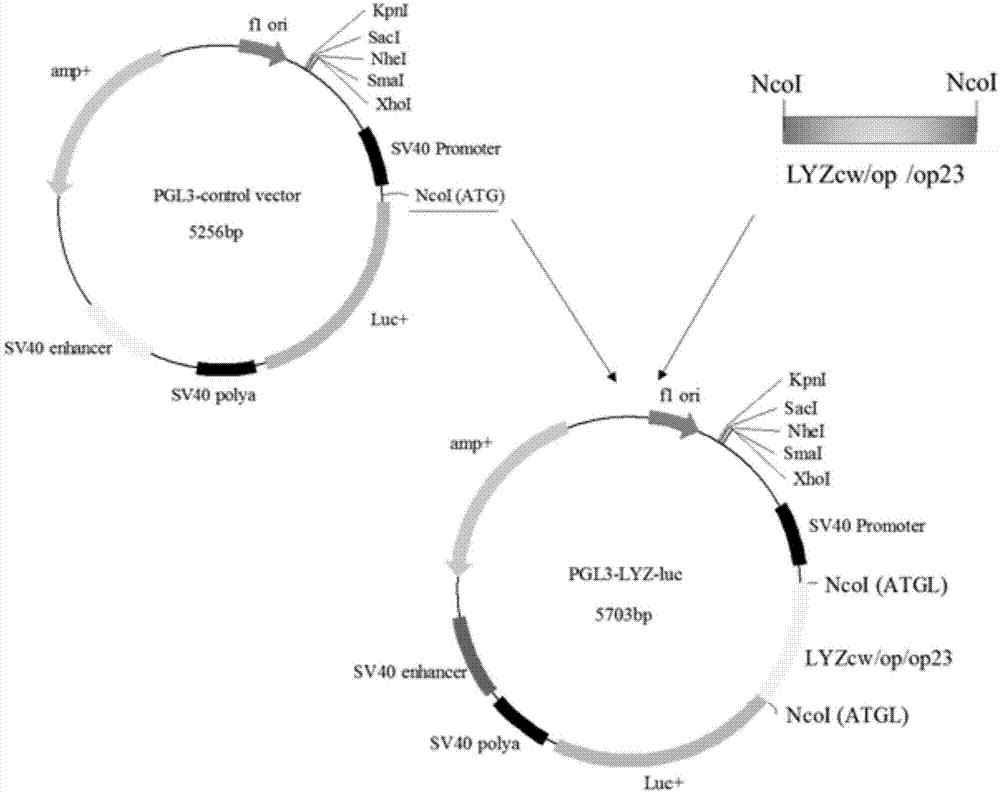
图1是PcDNA-Lyz(cw/op/op23)载体构建示意图;
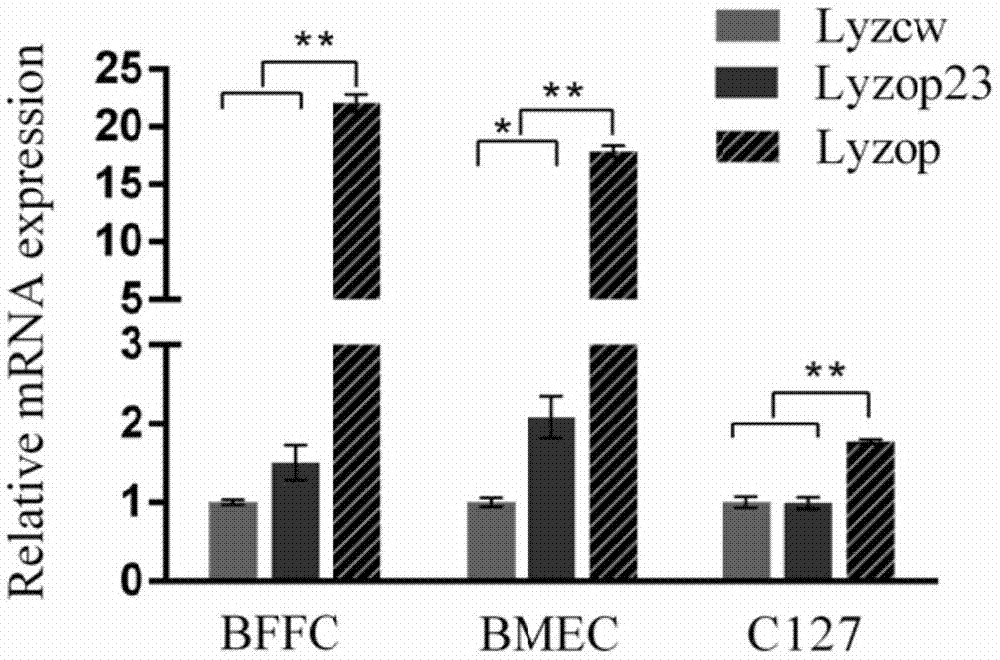
图2是qPCR比较人溶菌酶密码子优化mRNA水平表达量;
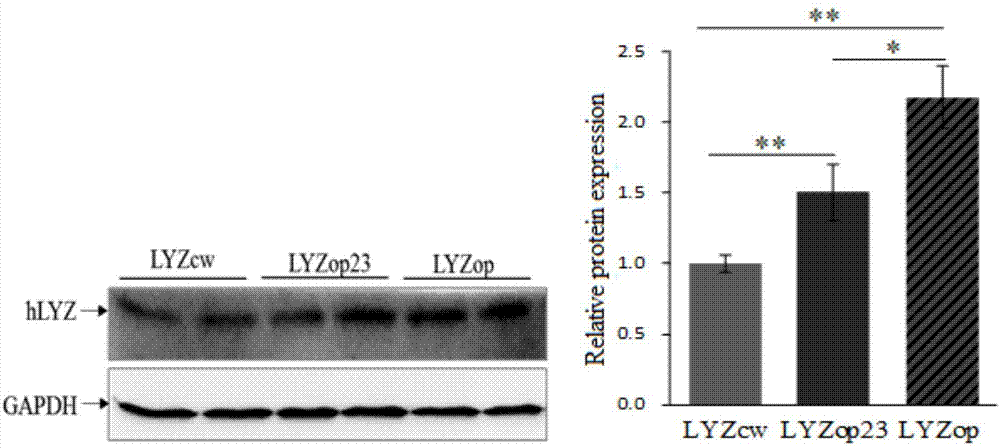
图3是Western-blot比较重组人溶菌酶密码子优化效果;
图4是PGL3-Lyz-luc载体构建示意图;
图5是PGL3-Lyz-luc载体NcoI酶切鉴定;
图6是荧光素酶评估密码子优化效果;
下面结合附图和实施例对本发明作进一步的详细说明。
具体实施方式
申请人通过比较发现,编码人溶菌酶基因密码子第三位碱基偏好以AT结尾,与其他真核生物如哺乳动物密码子,如牛乳腺中表达量高的乳蛋白编码基因密码子使用普遍偏好以CG结尾存在一定差异。经进一步研究表明,人溶菌酶与牛乳腺优势乳蛋白对简并密码子使用频率上存在明显差异,而这种差异可能是造成人溶菌酶在乳腺生物反应器中表达量低的主要原因。
申请人依据计算、分析获得的牛乳腺中乳蛋白高频密码子(使用频率较高) 及低频密码子(稀有密码子)对人溶菌酶密码子进行优化,提高人溶菌酶编码基因中对应氨基酸牛优势乳蛋白中高频密码子,减少低频密码子。比较发现,在不改变人溶菌酶蛋白质序列的基础上,经过密码子优化设计,可以大幅度提高重组人溶菌酶的表达量。同时,利用人溶菌酶基因与PGL3-control载体Nco1 酶切位点连接实现融合表达,可以通过检测荧光素酶表达量来比较密码子优化效果,相比实时荧光定量PCR及Western-blot等方法,进一步提高了密码子优化效果评价方法的准确度和便捷性。为利用细胞反应器及乳腺生物反应器生产重组人溶菌酶提供了基因载体,并为今后开展基因密码子优化效果分析提供了一种可靠且快速的评价方法。
通过对人溶菌酶基因密码子重新设计并优化,大幅度提高其体外表达水平,并通过人溶菌酶基因与荧光素酶融合表达,快速、便捷及准确的评估密码子优化效果的方法。
申请人利用CodonW和EMBOSS等软件对牛优势乳蛋白基因密码子使用偏好分析结果表明,牛乳蛋白基因密码子偏好以GC结尾,而人溶菌酶基因密码子偏好以AT结尾。进一步分析发现,牛乳蛋白基因对CTG(2.71)、AGG(2.32)、 GCC(1.73)、GTG(1.68)、ATC(1.64)等5个密码子具有明显的偏好性(同义密码子相对使用度RSCU>1.5),为高频密码子。而TTA(0.12)、ATA(0.18)、 TCG(0.19)、CTA(0.25)、CCG(0.26)、GCG(0.45)、ACG(0.48)、TAG(0.43)等8个密码子在牛乳蛋白基因中使用频率较低(RSCU<0.5),为低频密码子。根据牛乳蛋白高频密码子及低频密码子使用情况,对人溶菌酶密码子进行了优化。其中Lyzop对溶菌酶全部密码子进行了全局优化,而Lyzop23只对翻译起始后23位密码子进行了优化。通过CodonW比较优化前后序列结果表明, Lyzcw、Lyzop23、Lyzop第三位碱基GC3含量分别为0.407、0.450、0.757。RNA 二级结构最小自由能从-138.83kcal/mol分别提高到了-141.02kcal/mol和-178.12 kcal/mol。表明,密码子优化提高了人溶菌酶基因mRNA的稳定性。
通过构建Lyzcw-PcDNA、Lyzop-PcDNA及Lyzop23-PcDNA三个人溶菌酶表达质粒,分别转染牛胎儿成纤维细胞(BFFC)、牛乳腺上皮细胞(BMEC) 和小鼠乳腺上皮细胞(C127)(每种质粒转染3个重复)。在转染后24h收集RNA 进行人溶菌酶转录水平检测。定量结果表明,Lyzop23相比Lyzcw在BFFC和 BMEC细胞中分别提高了1.5倍(P>0.05)和2.08倍(P<0.05)。而Lyzop相比野生型lyzcw在上述两个细胞中分别提高了22倍(P<0.01)和17.8倍(P<0.01)。上述结果表明,根据牛乳腺优势蛋白进行密码子优化后的人溶菌酶能显著提高其在牛成纤维细胞及乳腺细胞中mRNA稳定性及转录水平表达量。对转染牛乳腺上皮细胞提取蛋白进行Western-blot结果表明,Lyzop和Lyzop23相比Lyzcw 分别提高了2.18倍(P<0.01)和1.5倍(P<0.01)。上述结果表明,根据牛乳腺优势蛋白进行密码子优化后的重组人溶菌酶基因能显著提高其在牛乳腺细胞中蛋白表达量。
进一步,将上述三个基因序列通过Nco1酶切位点克隆到PGL3-control载体上。构建好的载体通过酶切鉴定及测序验证。测序结果表明,Lyzcw、Lyzop23 和Lyzop编码区与萤火虫荧光素酶基因正确融合,没有移码突变。
通过荧光素酶检测结果表明,Lyzop相比Lyzcw在BFFC、BMEC、C127 细胞中分别提高了2.44、1.96和1.99倍,差异极显著(P<0.01)。Lyzop23密码子优化方式相比野生型Lyzcw在BMEC中提高了1.48倍,差异极显著(P<0.01)。
以下是发明人给出的具体实施例和具体操作过程,但本发明不限于下述的实施例。
以下实施例涉及的载体、菌株、细胞、试剂来源如下:
牛成纤维细胞为国家肉牛改良中心秦川牛耳尖皮肤组织通过酶消化法分离获得。
牛乳腺上皮细胞、小鼠C127细胞购自中科院上海细胞库,TOPO10感受态细菌、DH5α购自天根生化科技有限公司。
RT-PCR试剂盒购自Takara公司,质粒提取、胶回收试剂盒购自OMEGA 公司,双荧光素酶试剂盒购至Promega公司。
19-T、内切酶、连接酶、实时荧光定量PCR试剂盒均购自Takara公司,KOD-PLUS高保真PCR酶购自东洋纺公司。
DMEM、胎牛血清(FBS)均购自Hyclone公司。
Lipo2000转染试剂购至Invitrogen。
蛋白酶抑制剂购至Roche。
兔抗人溶菌酶多克隆抗体、兔抗GAPDH多克隆抗体(一抗)以及抗兔HRP 二抗均购至abcam公司。
其余Western-blot相关试剂均购置碧云天。
引物合成在南京金斯瑞公司和Invitrogen公司完成。
酶切、连接、回收、转化、RNA提取、PCR扩增等常规分子生物学实验操作步骤详见《分子克隆(第三版)》。
实施例1:牛优势乳蛋白密码子使用偏好分析
利用CodonW和EMBOSS等软件对牛优势乳蛋白基因密码子使用偏好分析结果表明,牛乳蛋白基因密码子偏好以GC结尾,而人溶菌酶基因密码子偏好以AT结尾。
本实施例中,申请人根据各种蛋白在乳腺中的含量,选取了αs1,αs2,β,K酪蛋白,β-乳球蛋白,乳清白蛋白,乳铁蛋白七种在乳腺中表达量较高的优势乳蛋白作为分析对象,通过查找NCBI获得上述蛋白编码基因CDS区序列。具体乳蛋白基因名称、GenBank登录号、CDS序列长度见附表1。
附表1:牛优势乳蛋白基因序列信息
对7个牛乳优势蛋白基因共59个密码子(不含起始密码子甲硫氨酸(M)) 进行密码子使用相对概率(RSCU)分析。RSCU>1.5则表示该密码子为高频密码子,而RSCU<0.5则表示为稀有密码子。通过EMBOSS中CUSP软件分析结果表明,牛乳腺中7个优势牛乳蛋白基因对CTG(2.71)、AGG(2.32)、GCC (1.73)、GTG(1.68)、ATC(1.64)等5个密码子具有明显的偏好性(RSCU>1.5,附表2,方框标注),为高频密码子。而TTA(0.12)、ATA(0.18)、TCG(0.19)、 CTA(0.25)、CCG(0.26)、GCG(0.45)、ACG(0.48)、TAG(0.43)等8个密码子在牛乳蛋白基因中为稀有密码子(RSCU<0.5,见附表2,下划线标注),为低频密码子。
附表2:牛乳蛋白密码子使用偏好性分析
注:方框表示牛乳蛋白基因中高频密码子,下划线表示牛乳蛋白基因中低频密码子。
上述结果表明,牛乳腺中,CTG、AGG、GCC、GTG、ATC为使用偏好密码子,而TTA、ATA、TCG、CTA、CCG、GCG、ACG、TAG为稀有密码子。
实施例2:人溶菌酶基因密码子优化设计
根据上述CodonW软件分析结果表明,人溶菌酶密码子第三位碱基偏好以 AT结尾(第三位碱基GC3s含量平均为0.41),而牛优势乳蛋白基因第三位密码子偏好以GC结尾(第三位碱基GC3s含量平均为0.54)。根据附表2牛乳蛋白高频密码子及低频密码子使用情况,发明人对人溶菌酶密码子进行了优化设计。其中lyzop对人溶菌酶基因全部密码子进行了优化,而Lyzop23只对人溶菌酶基因前面23个密码子进行了优化(附表3)。
附表3:人溶菌酶基因密码子优化情况
注:斜体加粗部分表示重组人溶菌酶基因按照牛乳蛋白偏好密码子进行设计优化后的密码子。
通过人溶菌酶密码子对RNA稳定性比较分析(附表4),CodonW比较优化前后序列结果表明,Lyzcw、Lyzop23、Lyzop第三位碱基GC含量分别为0.407、 0.450、0.757。RNA二级结构最小自由能从-138.83kcal/mol分别提高到了-141.02 kcal/mol和-178.12kcal/mol。表明,密码子优化提高了人溶菌酶基因mRNA的稳定性。
附表4:人溶菌酶密码子对RNA稳定性比较分析
实施例3:密码子优化设计对人溶菌酶mRNA转录水平影响检测
将Lyzcw、Lyzop23、Lyzop根据附表3中序列进行全基因合成,并添加 HindIII和XhoI酶切位点。
进一步,HindIII和XhoI分别酶切Lyzcw、Lyzop23、Lyzop以及PcDNA3.1 (+)载体,通过胶回收上述片段,并利用T4DNA酶分别连接构建PcDNA-Lyzcw、 PcDNA-Lyzop23及PcDNA-Lyzop三个人溶菌酶表达质粒(见图1)。
将上述三个质粒经转染DH5α、挑单克隆摇菌、提质粒、HindIII和XhoI 双酶切鉴定后,挑选构建成功的PcDNA-Lyzcw、PcDNA-Lyzop23及 PcDNA-Lyzop质粒用于细胞转染。
培养的牛胎儿成纤维细胞(BFFC)、牛乳腺上皮细胞(BMEC)、小鼠乳腺上皮细胞(C127)汇合度达到80%时,传代接种到12孔板用于细胞转染。将 PcDNA-Lyzcw、PcDNA-Lyzop23及PcDNA-Lyzop按照Lipo2000说明书(脂质体:质粒=3μl:1μg比例)分别转染BFFC、BMEC和C127细胞(每种质粒分别转染BFFC、BMEC和C127 3种细胞,且每种细胞设置3个重复)。24h后,通过Trizol法提取细胞RNA(DNA酶DNaseI处理,去除基因组DNA及残留质粒)。利用反转录试剂盒,分别取2μg RNA反转录获得cDNA。
利用ABI7500荧光定量PCR仪对不同转染组中重组人溶菌酶mRNA表达量进行分析。实时荧光定量PCR结果表明,Lyzop23相比Lyzcw在BFFC和BMEC细胞中分别提高了1.5倍(P>0.05)和2.08倍(P<0.05)。而Lyzop相比野生型Lyzcw在上述两个细胞中分别提高了22倍(P<0.01)和17.8倍(P <0.01)。而在小鼠乳腺细胞中,Lyzop23相比Lyzcw没有明显变化,而Lyzop 相比Lyzcw提高了1.82倍(P<0.01)(图2)。上述结果表明,根据牛乳腺优势蛋白进行密码子优化后的人溶菌酶基因能显著提高其在牛成纤维细胞、牛乳腺上皮细胞的转录水平mRNA表达量。
实施例4:密码子优化设计对人溶菌酶蛋白水平表达影响检测
如实施例3所述,PcDNA-Lyzcw、PcDNA-Lyzop23及PcDNA-Lyzop三个人溶菌酶表达载体转染牛BMEC细胞(每种细胞设置3个重复)。
具体的步骤是:转染48h后,经预冷PBS洗涤(2遍),然后IP裂解液裂解(200μl,冰上裂解30分钟),离心(15000转,10分钟),蛋白浓度测定,添加5×上样缓冲液煮沸(10分钟),即蛋白样品制备完成,用于Western-blot 检测。
制备15%PAGE蛋白电泳,分别取50μg Lyzcw、Lyzop23、Lyzop三个蛋白样本上样用于蛋白电泳。待电泳指示剂溴酚蓝跑到底部时终止电泳。取出电泳胶,按照伯乐半干转说明,200mA,1小时完成蛋白转膜。
然后经过浓度为5%的奶粉封闭1小时,经剪膜、一抗孵育1小时(1:1000 稀释)、TBST洗膜3次(10分钟/次)、二抗孵育1h(1:3000稀释)、TBST洗膜(10分钟/次)、显色成像等步骤,检测密码子优化对重组人溶菌酶蛋白表达影响。
结果表明,Lyzop和Lyzop23相比Lyzcw分别提高了2.18倍(P<0.01)和 1.5倍(P<0.01)(图3)。上述结果表明,根据牛乳腺优势蛋白进行密码子优化后的人溶菌酶基因能显著提高其在牛乳腺细胞中蛋白表达量。
实施例5:人溶菌酶基因与荧光素基因融合实现密码子优化效果快速评价
以PcDNA-Lyzcw、PcDNA-Lyzop23及PcDNA-Lyzop载体为模板,设计引物克隆获得两端带有NcoI酶切位点的Lyzcw、Lyzop23、Lyzop序列,其中反向引物序列去掉Lyzcw、Lyzop23、Lyzop序列中翻译终止密码子(TAA),通过 PGL3-control NcoI酶切位点与萤火虫荧光素酶基因实现融合翻译。通过NcoI 内切酶分别酶切上述三个序列片段和PGL3-control载体并胶回收酶切后片段。根据T4DNA连接酶说明分别将Lyzcw、Lyzop23、Lyzop片段与PGL3-control 载体连接,转化细菌并培养16h后挑细菌克隆摇菌,提质粒(图4)。构建好的 PGL3-Lyzcw、PGL3-Lyzop23、PGL3-Lyzop质粒经NcoI酶切鉴定和测序鉴定 (图5)。结果表明,Lyzcw、Lyzop23和Lyzop编码区与萤火虫荧光素酶基因正确融合,没有移码突变。
将构建好的PGL3-Lyzcw、PGL3-Lyzop23、PGL3-Lyzop质粒分别于Prl-tk 海肾萤火虫荧光素酶按照50(μg):1(μg)比例按照实施例3分别共转染BFFC、 BMEC和C127三种细胞(每种细胞3个重复),通过荧光素酶检测结果表明, Lyzop相比Lyzcw在BFFC、BMEC、C127细胞中分别提高了2.44、1.96和1.99 倍,差异极显著(P<0.01)。Lyzop23密码子优化方式相比野生型Lyzcw在BMEC 中提高了1.48倍,差异极显著(P<0.01)(图6)。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明技术原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
核苷酸或氨基酸序列表
<110>西北农林科技大学
<120>一种密码子优化的重组人溶菌酶基因及其应用
<160>
<210> 1
<211> 447
<212> Lyzcw序列
<213>核苷酸
<220>
<400>
ATGAAGGCTCTCATTGTTCTGGGGCTTGTCCTCCTTTCTGTTACGGTCCAGGGCAAGGTCTTTGAAAGGTGTGAGTTGGCCAGAACTCTGAAAAGATTGGGAATGGATGGCTACAGGGGAATCAGCCTAGCAAACTGGATGTGTTTGGCCAAATGGGAGAGTGGTTACAACACACGAGCTACAAACTACAATGCTGGAGACAGAAGCACTGATTATGGGATATTTCAGATCAATAGCCGCTACTGGTGTAATGATGGCAAAACCCCAGGAGCAGTTAATGCCTGTCATTTATCCTGCAGTGCTTTGCTGCAAGATAACATCGCTGATGCTGTAGCTTGTGCAAAGAGGGTTGTCCGTGATCCACAAGGCATTAGAGCATGGGTGGCATGGAGAAATCGTTGTCAAAACAGAGATGTCCGTCAGTATGTTCAAGGTTGTGGAGTGTAA
<210> 2
<211> 447
<212> Lyzop序列
<213>核苷酸
<220>
<400>
ATGAAGGCCCTGATCGTGCTGGGACTGGTGCTGCTGTCTGTGACCGTGCAGGGCAAGGTGTTTGAGAGGTGCGAGCTGGCCAGGACTCTGAAGAGGCTGGGAATGGATGGCTATAGGGGAATCAGCCTGGCCAACTGGATGTGTCTGGCCAAGTGGGAGTCTGGCTATAACACCAGGGCCACCAACTATAACGCCGGAGACAGGAGCACCGACTATGGCATCTTTCAGATCAACTCTAGGTATTGGTGTAACGACGGCAAGACCCCTGGCGCCGTGAATGCCTGTCACCTGTCTTGTAGCGCCCTGCTGCAGGACAACATCGCCGATGCTGTGGCCTGCGCCAAGAGGGTGGTGAGGGATCCTCAGGGAATCAGGGCTTGGGTGGCCTGGAGGAATAGGTGCCAGAACAGGGACGTGAGGCAGTATGTGCAGGGATGTGGAGTGTAA
<210> 3
<211> 447
<212> Lyzop23序列
<213>核苷酸
<220>
<400>
ATGAAGGCCCTGATCGTGCTGGGACTGGTGCTGCTGTCTGTGACCGTGCAGGGCAAGGTGTTTGAGAGGTGTGAGTTGGCCAGAACTCTGAAAAGATTGGGAATGGATGGCTACAGGGGAATCAGCCTAGCAAACTGGATGTGTTTGGCCAAATGGGAGAGTGGTTACAACACACGAGCTACAAACTACAATGCTGGAGACAGAAGCACTGATTATGGGATATTTCAGATCAATAGCCGCTACTGGTGTAATGATGGCAAAACCCCAGGAGCAGTTAATGCCTGTCATTTATCCTGCAGTGCTTTGCTGCAAGATAACATCGCTGATGCTGTAGCTTGTGCAAAGAGGGTTGTCCGTGATCCACAAGGCATTAGAGCATGGGTGGCATGGAGAAATCGTTGTCAAAACAGAGATGTCCGTCAGTATGTTCAAGGTTGTGGAGTGTAA
[收起]
附图
评论
全部评论
共{{commentCount}}条{{rs.Msg_Content}}